Εφαρμογή της Γενετικής
στη διερεύνηση της Κλινικής Αλλεργίας
ΑΠOΔOΣΗ ΣΤΑ ΕΛΛΗΝΙΚΑ: Φ. ΒΛΑΣΤOΣ
HIROHISA SAITO, ATSUSHI KATO & KENJI MATSUMOTO
Allergy Clin Immunol Int 2003; 15(5)
Εισαγωγή
Χρησιμοποιώντας τη γονιδιακή αλληλουχία [1-3] αναμένεται ότι θα απαντήσουμε
σε προηγουμένως αναπάντητες ερωτήσεις, όπως είναι η πιθανότητα εμφάνισης μιας
νόσου. Διαθέτουμε πλέον εργαλεία για τη διερεύνηση του ρόλου των ποικιλιών του
γονιδιακού DNA μεταξύ των ατόμων, όπως οι μεμονωμένοι πολυμορφισμοί (single
nucleotide polymorphism-SNP) στην παθογένεια νόσων και αποκρίσεων σε φάρμακα.
Η κατανόηση του γονιδιώματος θα βοηθήσει επίσης στην κατανόηση του μεταγραφώματος
[5], του συνόλου των μεταγραφών μέσα σε ένα κύτταρο και του πρωτεϊνώματος [6],
του συνόλου των πρωτεϊνών που ελέγχουν τη λειτουργία ενός κυττάρου (σχήμα 1).
Μέχρι πρόσφατα, χρειαζόταν πολύς κόπος και χρόνος προκειμένου να μετρηθεί η
έκφραση γονιδίων. Ωστόσο, με τη χρήση της προσφάτως ανακαλυφθείσης τεχνολογίας
των chips DNA (cDNA microarrays, high-density oligonucleotide probe arrays)
[7,8], αυτή η συστηματική ανάλυση των μεταγραφωμάτων έγινε εύκολη. Κατά τα τελευταία
χρόνια, αυτή η τεχνολογία εξαπλώθηκε σε πολλές περιοχές της κλινικής ιατρικής
[7-10], συμπεριλαμβανομένης της δικής μας [11-19]. Εδώ παρουσιάζεται η ερευνητική
στρατηγική των συγγραφέων στην ευρεία ανάλυση του γονιδιώματος των SNP σχετικά
με το άσθμα.
Στρατηγική για
την ανακάλυψη των SNP
που είναι υπεύθυνες για το άσθμα
Σε αντίθεση με
μια «μετάλλαξη» που υπάρχει σε <1% του συνόλου του πληθυσμού και συχνά προκαλούν
σοβαρές κληρονομικές παθήσεις, οι SNP εμφανίζονται σε >1% του πληθυσμού και
καθορίζουν κοινά ατομικά χαρακτηριστικά, όπως το χρώμα των τριχών [4,2]. Μεμονωμένοι
πολυμορφισμοί νουκλεοτιδίων εμφανίζονται στο 0.1% όλων των αλληλουχιών του DNA
και είναι οι συχνότερες γενετικές διαφορές μεταξύ ατόμων. Συνεπώς, ελέγχοντας
τους SNP, υπολογίζεται ότι μπορεί να καθοριστεί η έναρξη νόσων, όπως η αρτηριακή
υπέρταση [21] ή η ευαισθησία σε φάρμακα [22]. Μετάλλαξη που οφείλεται σε μεμονωμένη
αντικατάσταση νουκλεοτιδίου στην περίπτωση επίκτητης νόσου και ο SNP που είναι
υπεύθυνος για την ευαισθησία σε φάρμακα, συνοδεύονται συχνά από ολική απώλεια
της λειτουργίας της αντίστοιχης πρωτεΐνης. Αντίθετα, οι SNP που είναι υπεύθυνοι
για κοινές ασθένειες συνοδεύονται από ήπια μείωση του επιπέδου έκφρασης ή της
λειτουργίας προϊόντων των γονιδίων και θα μπορούσαν να επηρεάζονται από περιβαλλοντικούς
παράγοντες. Από τις πολυάριθμες νουκλεοτιδικές αντικαταστάσεις που είναι υπεύθυνες
για την ευαισθησία σε φάρμακα, σε περιπτώσεις όπου το προϊόν του γονιδίου δεν
είναι απαραίτητο για την επιβίωση του ανθρώπου, η ποικιλία είναι αρκούντως συχνή
[23]. Όμως, όταν το προϊόν του γονιδίου είναι ουσιώδες για την επιβίωση, η λειτουργική
απώλεια θα πρέπει να είναι μικρή.
Το βρογχικό άσθμα θεωρείται συχνά ως ατοπική νόσος, αλλά στην πραγματικότητα
είναι μια ομάδα παθήσεων με παρόμοια συμπτώματα, με πολλούς πυροδοτικούς παράγοντες
[24]. Για παράδειγμα, το παιδικό και το άσθμα των ενηλίκων έχουν πολλές διαφορές
ως προς τους πυροδοτικούς παράγοντες.
Υπάρχουν πολλοί ενήλικες ασθενείς με άσθμα, αλλά όχι παιδιά, η νόσος των οποίων
σχετίζεται με φλεγμονή ανεξάρτητη από τις ανοσοσφαιρίνες IgE και παθολογική
αναδιαμόρφωση των αεραγωγών, ως συνέπεια της φλεγμονής [25] ή της ευαισθησίας
στην ασπιρίνη [2]. Αντίθετα, οι λοιμώξεις με ρινοϊούς είναι συχνοί πυροδότες
των κρίσεων του παιδικού άσθματος [27]. Συνεπώς, για την ανίχνευση των SNP σε
ένα μόριο που επηρεάζει έναν ορισμένο τύπο άσθματος, πρέπει να προσδιορίσουμε
τον τύπο του συγκεκριμένου άσθματος, αναλύοντας την κλινική του συμπεριφορά.
 |
 |
|
Σχήμα 1. Σχηματική απεικόνιση του γονιδιώματος του μεταγραφώματος και του πρωτεϊνώματος. |
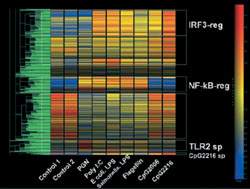
Σχήμα 2. Ενα παράδειγμα ιεραρχικής ανάλυσης για την ανίχνευση των gene clusters. |
Δοκιμασίες
μεταγραφώματος για την ανίχνευση νέων διαγνωστικών μορίων
Για την κατανόηση των λειτουργικών ρόλων του συνόλου του γονιδιώματος, ίσως
να είναι απαραίτητη μια ανάλυση του πρωτεϊνώματος. Ωστόσο, η ανάλυση του πρωτεϊνώματος
δεν είναι ακόμη ευχερής, αν και οι πρόσφατες τεχνολογικές πρόοδοι έκαναν εφικτή
την άμεση μέτρηση μερικών εκατοντάδων πρωτεϊνών. Πράγματι, η πλειοψηφία των
πρωτεϊνών που υπάρχουν στο πρωτεΐνωμα βρίσκονται ακόμη κρυμμένες στο «μαύρο
κουτί» και ο αριθμός τους υπολογίζεται περίπου >100.000, χωρίς να μετρώνται
διάφορες ανοσοσφαιρίνες και υποδοχείς των Τ λεμφοκυττάρων. Συνεπώς, παραμένει
κοπιώδης η μέτρηση ακόμη και του 1% των πρωτεϊνών του πρωτεϊνώματος. Σε αντίθεση
με τα πρωτεϊνώματα, η επεξεργασία του μεταγραφώματος έγινε ήδη εφικτή μετά από
τις προόδους στην τεχνολογία του microarray. Σήμερα, υπάρχουν διαθέσιμα εργαλεία
για την ταυτόχρονη μέτρηση των επιπέδων έκφρασης 30.000 γονιδίων. Σχεδόν το
σύνολο της έκφρασης όλων των γονιδίων μπορεί να μετρηθεί με την τεχνολογία των
chip του DNA, εφόσον ο αριθμός των γονιδίων που υπάρχουν στο γονιδίωμα υπολογίζεται
σε 32.000. Το «μεταγράφωμα» (transcriptome) αφορά όλους, όχι μερικούς, μεταγραφικούς
παράγοντες που εκφράζονται σε ένα κύτταρο. Δεν πρόκειται να μας διαφεύγουν πλέον
τυχόν ανεπιθύμητες ενέργειες νέων φαρμάκων, εφόσον διαθέτουμε μια τέτοια τεχνολογία.
Η
ανίχνευση γονιδίων που είναι υπεύθυνα για την πρόκληση αλλεργικών παθήσεων
Ο όρος «ατοπία» προτάθηκε το 1923. Αρχικά, η «ατοπία» κατανοήθηκε ως μια κληρονομούμενη
προδιάθεση σε υπερευαισθησία άμεσου τύπου σε κοινά αλλεργιογόνα. Ωστόσο, η ατοπία
χρησιμοποιείται σήμερα για να υποδηλώσει «μηχανισμούς διαμεσολαβούμενους από
τις IgE», που μπορεί να είναι και επίκτητοι, ως αποτέλεσμα περιβαλλοντικών επιδράσεων.
Όταν η ατοπία καθορίζεται ως μία ή δύο θετικές αντιδράσεις για ειδικά IgE αντισώματα
έναντι κοινών αλλεργιογόνων in vitro ή χρησιμοποιώντας τις δερματικές δοκιμασίες,
ο επιπολασμός της ατοπίας αυξάνεται.
Υπάρχει μια γενικευμένη αύξηση στην ευαισθητοποίηση μέσω ανοσοσφαιρινών IgE.
Τα παιδιά που μεγαλώνουν στην ύπαιθρο έχουν μια μειωμένη τάση για ατοπικές παθήσεις
[30,31]. Αν και ο ακριβής μηχανισμός παραμένει αδιευκρίνιστος, πρόσφατες μελέτες
υποδεικνύουν ότι η έκθεση στην ενδοτοξίνη και άλλα σε μικροβιακά παράγωγα κατά
την παιδική ηλικία, ίσως παίζει σημαντικό ρόλο στη μειωμένη προδιάθεση των ατόμων
σε αλλεργικές παθήσεις [32,33]. Συνεπώς, η διαφορά στον επιπολασμό της ατοπίας
μεταξύ νέων και ηλικιωμένων μπορεί να οφείλεται στην ταχεία ανάπτυξη των συνθηκών
υγιεινής, σε χώρες όπως η Ιαπωνία κατά τη δεκαετία του 1950. Σύμφωνα με ερευνητές,
η υγιεινή και η εκδυτικοποίηση της διατροφής ίσως διευκολύνουν την εμφάνιση
ατοπίας, επηρεάζοντας τους μικροοργανισμούς που αλληλεπιδρούν με το λεμφικό
ιστό του εντέρου. Οι Shirakara και συνεργάτες ανέφεραν αντίστροφη σχέση μεταξύ
της απόκρισης στη φυματίνη και των επιπέδων της IgE, και υπέθεσαν ότι ίσως το
μυκοβακτηρίδιο της φυματίωσης μπορεί να τροποποιεί την ανοσολογική απάντηση
προς την κατεύθυνση του τύπου Th-135. Αρκετές μεταγενέστερες μελέτες επιβεβαίωσαν
ότι το μυκοβακτηρίδιο της φυματίωσης ή τα συστατικά του [36,37,38] έχουν ένα
προληπτικό αποτέλεσμα ως προς την εμφάνιση αλλεργικών παθήσεων, αν και το αποτέλεσμα
αυτό περιορίζεται στα πρώτα χρόνια του βίου. Από αυτές τις μελέτες, μπορεί να
υποθέσει κανείς ότι τα μικρόβια ή τα συστατικά τους, όπως η ενδοτοξίνη, μπορεί
να μεταβάλουν την ανοσολογική απάντηση που σχετίζεται με την εμφάνιση αλλεργικών
παθήσεων κατά την παιδική ηλικία, ιδιαίτερα εάν τα μικρόβια εντοπίζονται στα
άκρα του πεπτικού σωλήνα ή στην τροφή.
Ποιοί είναι οι μηχανισμοί που συμμετέχουν στην επίδραση των μικροοργανισμών
στην ανοσολογική απάντηση νωρίς κατά τη διάρκεια του βίου; Πρόσφατα, η εγγενής
ανοσολογική κατάσταση τέθηκε στο στόχαστρο των ερευνητών, όχι μόνο επειδή συμμετέχει
στην πρόληψη των μικροβιακών λοιμώξεων των διαφόρων οργάνων, αλλά επίσης επειδή
είναι ικανή να μετατρέψει την ανοσολογική απάντηση του ξενιστή από την αλλεργικού
τύπου απάντηση (Th2), στην απάντηση μη αλλεργικού τύπου (Th1) [39,40]. Μόρια
που διατηρούνται μεταξύ των ειδών εμπλέκονται σε αυτές τις διαδικασίες ως ειδικοί
υποδοχείς [39]. Τα μόρια που είναι υπεύθυνα για την εγγενή ανοσία καλούνται
Toll-like receptors (TLRs)41, είναι ομόλογα των Toll της Drosophila [42], που
αναγνωρίζουν μοριακά πρότυπα σχετικά με παθογόνα μικρόβια [39]. Σήμερα [40],
έχουν ήδη ταυτοποιηθεί ανθρώπινοι TLRs [43]. Συνεπώς, για την πρόληψη των αλλεργικών
παθήσεων κατά την παιδική ηλικία, αναμένεται να αποδειχθεί αποτελεσματική η
διέγερση μέσω ουσιών που συνδέουν τους TLRs, π.χ. από μικροβιακά παράγωγα. Ωστόσο,
ορισμένες ουσίες που συνδέονται με τους TLRs, όπως η ενδοτοξίνη, προκαλούν σοβαρή
ιστική φλεγμονή, γεγονός που απειλεί τους ιστούς με χρόνιες βλάβες. Σε μια προκαταρκτική
μελέτη, εξετάσαμε μεταγραφώματα από ανθρώπινα μονοκύτταρα υγιών εθελοντών, τα
οποία είχαμε διεγείρει με διάφορες ουσίες που συνδέονται με τους TLRs, κυρίως
μικροβιακά παράγωγα. Όπως θα ανέμενε κανείς, στους ανθρώπους, η ουσία που συνδέεται
με τους υποδοχείς TLR9 CpG44 προκάλεσε την έκφραση πολλών γονιδίων που σχετίζονται
με την αντι-ιική άμυνα και ιδιαίτερα την επιστράτευση των κυττάρων τύπου Th1,
χωρίς να προκαλεί την έκφραση των γονιδίων των προφλεγμονωδών κυτταροκινών,
όπως οι TNF-a και COX-2 (σχήμα 2). Πρόσφατα, πολλοί ερευνητές ανακοίνωσαν την
ταυτόχρονη αύξηση τόσο σε αλλεργικές παθήσεις που διαμεσολαβούνται από αντιδράσεις
τύπου Th2, όσο και αυτοάνοσων διαταραχών τύπου Th1, στη Δυτική Ευρώπη [45-47].
Σε αυτές τις αναφορές, περιλαμβάνονται και σχόλια σχετικά με τα πρόσφατα ευρήματα
των ρυθμιστικών κυτταρικών τύπων Th1 και Th 248-50. Στο προκαταρκτικό μας πείραμα,
η CpG προκάλεσε την παραγωγή μικρότερων επιπέδων γονιδίων CD25, IL-10, TGF-β
συγκριτικά με άλλες ουσίες που συνδέονται με τους υποδοχείς TLR. Συνεπώς, η
ανάλυση του ανοσολογικού μεταγραφώματος ή του πρωτεϊνώματος που επάγεται από
τους TLRs, μπορεί να αποδειχθεί χρήσιμη στο εγγύς μέλλον, πριν από την εφαρμογή
εξατομικευμένων μιγμάτων ουσιών που συνδέονται με τους TLRs, σε παιδιά υψηλού
κινδύνου για αλλεργικές παθήσεις.
Αναλύσεις του
μεταγραφώματος για την ανίχνευση μη αναμενόμενων περιστατικών
Η ανάλυση του μεταγραφώματος είναι επίσης χρήσιμη για την ανίχνευση μη αναμενόμενων
περιστατικών, που συχνά είναι αντίθετα ή μη σχετιζόμενα με δημοσιευμένα στοιχεία.
Ερευνήσαμε την έκφραση ειδικών για ορισμένα κύτταρα γονιδίων στο μεταγράφωμα
ανθρώπινων μαστοκυττάρων και συγκρίναμε με άλλα κύτταρα. Σε σύγκριση με τις
αναμενόμενες πρωτεΐνες των μαστοκυττάρων, όπως οι τρυπτάσες, η μείζων βασική
πρωτεΐνη, που εθεωρείτο μια πρωτεΐνη ειδική των ηωσινοφίλων, βρέθηκε να εκφράζεται
σε μεγάλο βαθμό στα μαστοκύτταρα [12]. Μαστοκύτταρα από τέσσερις διαφορετικές
πηγές βρέθηκαν να περιέχουν υψηλά επίπεδα μείζονος βασικής πρωτεΐνης στα κοκκία
τους. Χωρίς τη χρήση των chips DNA, κανείς δεν θα είχε ανακαλύψει ένα τόσο αναπάντεχο
εύρημα. Τα μοντέλα πειραματοζώων με αλλεργική φλεγμονή υπήρξαν ιδιαίτερα χρήσιμα.
Η χρήση μοντέλων ποντικών στο άσθμα αυξήθηκε κυρίως χάρη στη δυνατότητα της
επιλογής με την μέθοδο knock out των γονιδίων που αναστέλλουν συγκεκριμένες
παθοφυσιολογικές οδούς, οι οποίες εμπλέκονται στην παθοφυσιολογία της νόσου.
Ωστόσο, υπάρχει διχογνωμία ως προς την αξιοπιστία αυτών των μοντέλων του άσθματος
[51,52]. Πρέπει να σημειωθεί ότι τα μοντέλα ποντικών δεν εμφανίζουν τις επιθηλιακές
βλάβες που οφείλονται στην εναπόθεση συστατικών από τα ηωσινόφιλα κοκκία, όπως
η μείζων βασική πρωτεΐνη [51].
Έτσι, χρησιμοποιήσαμε GeneChip για να συγκρίνουμε τα μεταγραφώματα ανθρώπινων
και ζωικών μαστοκυττάρων, επειδή τα μαστοκύτταρα των τρωκτικών είναι κοινά εργαλεία
έρευνας, αλλά είναι αρκετά διαφορετικά από τα ανθρώπινα αντίστοιχα κύτταρα,
ως προς την απόκρισή τους σε ορισμένες κυτταροκίνες και φάρμακα.
Μετά από διέγερση μέσω υποδοχέων IgE με υψηλή συγγένεια (FcεRI), τα μεταγραφικά
επίπεδα αρκετών χυμοκινών CC (Ι-309, φλεγμονώδης πρωτεΐνη των μακροφάγων MIP-1a
και MIP-Iβ) βρέθηκαν να είναι σημαντικά αυξημένα, τόσο στα μεταγραφώματα των
ανθρώπινων μαστοκυττάρων, όσο και σε αυτά των μαστοκυττάρων των ποντικών. Αυτά
τα αποτελέσματα υποδηλώνουν ότι τα μαστοκύτταρα παίζουν κρίσιμο ρόλο για την
επιστράτευση CCR-κυττάρων στους ιστούς κατά τρόπο εξαρτώμενο από τις IgE και
ότι η, μέσω των υποδοχέων FcεRI, επαγωγή πολλών χυμοκινών CC διατηρείται σε
μεγάλο βαθμό από τον άνθρωπο στον ποντικό. Αντίθετα, πολλά άλλα γονίδια εκφράζονται
επιλεκτικά σε κάθε είδος μαστοκυττάρων [13].
Οι μελέτες σχετικά με τη λειτουργία μορίων που εκφράζονται ιδιαίτερα μόνο στα
κύτταρα των ποντικών, πρέπει να ερμηνεύονται με προσοχή ως προς τις πιθανές
επεκτάσεις τους στον άνθρωπο. Εξάλλου, συγκριτικές μελέτες έκφρασης των γονιδιωμάτων
μεταξύ των ζωικών ειδών, θα μπορούσαν να φανούν ιδιαίτερα χρήσιμες για την προβολή
των πειραματικών δεδομένων από μελέτες σε πειραματόζωα, σε ό,τι αφορά την παθογένεση
των ανθρωπίνων νόσων.
Στο εγγύς μέλλον, η πιθανότητα του άσθματος, καθώς και των άλλων αλλεργικών
παθήσεων, θα εξετάζεται σε κάθε νοσοκομείο ελέγχοντας όλους τους SNP και τις
πληροφορίες από το μεταγράφωμα που σχετίζονται με αυτές τις νόσους.
Bιβλιογραφία
1. Venter JC, Adams MD, Myers EW et al. The sequence of the human genome. Science
2001; 291:1304-51.
2. International Human Genome Sequencing Consortium. Initial sequencing and
analysis of the human genome. Nature 2001; 409:860-921.
3. Waterston RH, Lindblad-Toh K, Birney E et al. Initial sequencing and comparative
analysis of the mouse genome. Nature 2002; 420:520-62.
4. Cargill M, Altshuler D, Ireland J et al. Characterization of single-nucleotide
polymorphisms in coding regions of human genes. Nat Genet 1999; 22:231-8.
5. Velculescu VE, Madden SL, Zhang L et al. Analysis of human transcriptomes.
Nat Genet 1999; 23:387-3.
6. Apweiler R, Biswas M, Fleischmann W et al. Proteome analysis database: Online
application of InterPro and CluSTr for the functional classification of proteins
in whole genomes. Nucleic Acids Res 2001; 29:44-8.
7. Duggan DJ, Bittner M, Chen Y, Meltzer P, Trent JM. Expression profiling using
cDNA microarrays. Nat Genet 1999; 21:10-14.
8. Lipshutz RJ, Fodor SPA, Gingeras TR, Lockhart DJ. High density synthetic
oligonucleotide arrays. Nat Genet 1999; 21:20-4.
9. Fambrough D, McClure K, Kazlauskas A, Lander ES. Diverse signaling pathways
activated by growth factor receptors induce broadly overlapping, rather than
independent, sets of genes. Cell 1999; 97:727-41.
10. Alizadeh AA, Eisen MB, Davis RE et al. Distinct types of diffuse large B-cell
lymphoma identified by gene expression profiling. Nature 2000; 403: 503-11
11. Iida M, Matsumoto K, Tomita H et al. Selective down-regulation of high affinity
IgE receptor (Fc epsilon RI) alpha-chain messenger RNA among transcriptome in
cord blood-derived versus adult peripheral blood-derived cultured human mast
cells. Blood 2001; 97:1016-22.
12. Nakajima T, Matsumoto K, Suto H et al. Gene expression screening of human
mast cells and eosinophils using high-density oligonucleotide probe arrays:
Abundant expression of major basic protein in mast cells. Blood 2001; 98: 1127-34.
13. Nakajima T, Inagaki N, Tanaka H et al. Marked increase in CC chemokine gene
expression in both human and mouse mast cell transcriptomes following Fc epsilon
receptor I cross-linking: An interspecies comparison. Blood 2002; 100:3861-8.
14. Heishi M, Kagaya S, KatsunumaT et al. High-density oligonucleotide array
analysis of mRNA transcripts in severe atopic dermatitis patients' peripheral
blood cells. Int Arch Allergy Immunol 2002; 129:57-66.
15. Seki Y, Inove H, Nagata N, et al. Suppressor of cytokine signaling 3 (SoCS3)
regulates onset and maintenance of type 2 helper T cell mediated allergic responses.
Nat Med 2003; DOI: 10.1038/Nm 896.
16. Matsumoto Y, Oshida T, Obayashi I et al. Identification of highly expressed
genes in peripheral blood T cells from patients with atopic dermatitis. Int
Arch Allergy Immunol 2002; 129:327-40.
17. Ogawa K, Hashida R, Miyagawa M et al. Analysis of gene expression in peripheral
blood eosinophils from patients with atopic dermatitis and in vitro cytokine-stimulated
blood eosinophils. Clin Exp Immunol 2003 (in press).
18. Saito H, Nakajima T, Matsumoto K. Human mast cell transcriptome project.
Int Arch Allergy Immunol 2001; 125:1-8.
19. Matsumoto K, Saito H. Role of eosinophils in asthma: Sarastro or the Queen
of the Night? Int Arch Allergy Immunol 2001; 125:290-6.
20. Bentley DR. The Human Genome Project: An overview. Med Res Rev 2000; 20:189-96.
21. Martin ER, Lai EH, Gilbert JR et al. SNPing away at complex diseases: Analysis
of single-nucleotide polymorphisms around APOE in Alzheimer disease. Am J Hum
Genet 2000; 67:383-94.
22. Sesti F, Abbott GW, Wei J et al. A common polymorphism associated with antibiotic-induced
cardiac arrhythmia. Proc Natl Acad Sci USA 2000; 97:10613-18.
23. Koch HG, McClay J, Loh EW et al. Allele association studies with SSR and
SNP markers at known physical distances within a 1 Mb region embracing the ALDH2
locus in the Japanese, demonstrates linkage disequilibrium extending up to 400kb.
Hum Mol Genet 2000; 9:2993-9.
24. Pride NB. Asthma. Definition and clinical spectrum. Br Med Bull 1992; 48:1-9.
25. Jeffery PK. Remodeling in asthma and chronic obstructive lung disease. Am
JRespir Crit Care Med 2001; 164:828-38.
26. Sousa AR, Parikh A, Scadding G, Corrigan CJ, Lee TH. Leukotriene-receptor
expression on nasal mucosal inflammatory cells in aspirin-sensitive rhinosinusitis.
N Engl J Med 2002; 347:1493-9.
27. Gem JE, Busse WW. Relationship of viral infections to wheezing illnesses
and asthma. Nat Rev Immunol 2002; 2:132-9.
28. Nakagomi T, Itaya H, Tominaga T, Yamaki M, Hisamatsu S, Nakagomi O. Is atopy
increasing? Lancet 1994; 343:121-2.
29. Saito H: Translation of human genome into clinical allergy. Allergol Int
2003;52(2):45-50.
30. von Mutius E. Environmental factors influencing the development and progression
of pediatric asthma. J Allergy Clin Immunol 2002; 109:8525-32.
31. Braun Fahrlander C, Gassner M, Grize L et al. Prevalence of hay fever and
allergic sensitisation in farmer's children and their peers living in the same
rural community. SCARPOL team. Swiss Study on Childhood Allergy and Respiratory
Symptoms with Respect to Air Pollution. Clin Exp Allergy 1999; 32:28-34.
32. Gereda JE, Leung DYM, Thatayatikom A et al. Relation between housedust endotoxin
exposure, type 1 T cell development, and allergen sensitisation in infants at
high risk of asthma. Lancet 2000; 32:1680-3.
33. Matricardi PM, Rosmini F, Rapicetta M, Gasbarrini G, Stroffolini T. Atopy,
hygiene, and anthroposophic lifestyle. San Marino Study Group. Lancet 1999;
354:430.
34. Matricardi PM, Rosmini F, Riondino S, Fortini M, Ferrigno L, Rapicetta M,
Bonini S. Exposure to foodborne and orofecal microbes versus airborne viruses
in relation to atopy and allergic asthma: Epidemiological study. BMJ 2000;320:412-417.
35. ShirakawaT, EnomotoT, Shimazu S, Hopkin JM. The inverse association between
tuberculin responses and atopic disorder. Science 1997; 275:77-79.
36. von Hertzen L, Klaukka T, Mattila H, Haahtela T. Mycobacterium tuberculosis
infection and the subsequent development of asthma and allergic conditions.
J Allergy Clin Immunol l999; 104:1211-1214.
37. Aaby P, Shaheen SO, Heyes CB, Goudiaby A, Hall AJ, Shiell AW, Jensen H,
Marchant A. Early BCG vaccination and reduction in atopy in Guinea-Bissau. Clin
Exp Allergy 2000; 30:644-650.
38. Hylkema MM, Timens W, Luinge M, Van Der Werf N, Hoekstra MO. The effect
of bacillus Calmette-Guerin immunization depends on the genetic predisposition
to Th2-type responsiveness. Am J Respir Cell Mol Biol. 2002; 27:244-249.
39. Medzhitov R, Janeway CA Jr. Innate immunity: The virtues of a non-clonal
system of recognition. Cell 1997; 91:295-8.
40. Krieg AM. The role of CpG motifs in innate immunity. Curr Opin Immunol 2000;
12:35-43.
41. Rock FL, Hardiman G, Timans JC, Kastelein RA, Bazan JF. A family of human
receptors structurally related to Drosophila Toll. Proc Natl Acad Sci USA 1998;
95:588-93.
42. Lemaitre B, Nicolas E, Michaut L, Reichhart JM, Hoffmann JA. The dorsoventral
regulatory gene cassette spatzle/Toll/cactus controls the potent antifungal
response in Drosophila adults. Cell 1996; 86:973-83.
43. Akira S, Takeda K, Kaisho T. Toll-like receptors: Critical proteins linking
innate and acquired immunity. Nat Immunol 2001; 2:675-80.
44. Hemmi H, Takeuchi O, Kawai T, Kaisho T, Sato S, Sanjo H, Matsumoto M, Hoshino
K, Wagner H, Takeda K, Akira S. A Toll-like receptor recognizes bacterial DNA.
Nature 2000; 408:740-745.
45. Stene LC, Nafstad P. Relation between occurrence of type 1 diabetes and
asthma. Lancet 2001 Feb 24; 357(9256):607-8.
46. Kero J, Gissler M, Hemminki E, Isolauri E. Could TH1 and TH2 diseases coexist?
Evaluation of asthma incidence in children with coeliac disease, type 1 diabetes,
or rheumatoid arthritis: a register study. J Allergy Clin Immunol 2001 Nov;108(5):781-3.
47. Sheikh A, Smeeth L, Hubbard R. There is no evidence of an inverse re-lationship
between TH2-mediated atopy and Th1 -mediated autoimmune disorders: Lack of support
for the hygiene hypothesis. J Allergy Clin Immunol 2003.
48. Levings MK, Sangregorio R, Roncarolo MG. Human CD25(+)CD4(+) T regulatory
cells suppress naive and memory T cell proliferation and can be expanded in
vitro without loss of function. J Exp Med 2001 Jun 4; 193(11):1295-302
49. Levings MK, Bacchetta R, Schulz U, Roncarolo MG. The role of IL-10 and TGF-beta
in the differentiation and effector function of T regulatory cells. Int Arch
Allergy Immunol Dec 2002; 129(4):263-76.
50. Sakaguchi S. Control of immune responses by naturally arising CD4+ regulatory
T cells that express toll-like receptors. J Exp Med 2003 Feb 17; 197(4):397-401.
51. Gelfand EW. Mice are a good model of human airway disease. Am J Respir Crit
Care Med 2002; 166:5-6.
52. Persson CGA. Mice are not a good model of human airway disease. Am J Respir
Crit Care Med 2002; 166:6-7.